Статья
Беспалов Б.И. Сферическая нейросетевая модель познавательного действия// Вестник Московского университета.Серия 14.Психология.- 2014.- №4 -с.56-75.
Автор(ы): Беспалов Борис Иванович
Аннотация
На основе идей Е.Н. Соколова о механизмах «векторного кодирования» в мозге информации об объекте построена сферическая нейросетевая модель зрительного познавательного действия человека, в которой учтены и выражены в математической форме некоторые нейрофизиологические данные о строении и функционировании реальных нейронных сетей мозга. В работах Е.Н. Соколова и его последователей в математической форме описывалась преимущественно работа предетекторов и детекторов нейронной сети. Новизна данной работы состоит в том, что в ней в математической форме и с помощью общих принципов векторного кодирования информации в мозге описаны основные стадии опознавательного действия, начиная от воздействия объекта на сетчатку глаза и заканчивая стадией выполнения ответа на стимул. Рассмотрена также психофизиологическая проблема, связанная с описанием перехода q-го значения свойства объекта из его физиологического состояния Fq на выходе предетекторов в неосознанно ощущаемое (психологическое) состояние Dq на выходе ансамбля детекторов, а затем в частично осознанное состояние Ψq на выходе нейронов памяти, генерирующих опознавательные эталоны и пр. Механизм выбора двигательного ответа на q-е значение свойства объекта соотносится в модели с декодированием («редукцией») многомерного вектора Рq, описывающего состояние объекта на выходе ансамбля мотонейронов. Разработанная нейросетевая модель познавательного действия позволяет объединить в рамках деятельностной психологии идеи Е.Н. Соколова о «векторном кодировании» в мозге свойств объекта и представления А.Н. Леонтьева и А.В. Запорожца о механизмах «активного психического отражения объекта субъектом».
PDF: /pdf/vestnik_2014_4/vestnik_2014-4_56-75.pdf
Поступила: 06.06.2014
Страницы: 56-75
Ключевые слова: сферическая нейросетевая модель; векторное кодирование информации в мозге; отражение объекта субъектом; психофизиологическая проблема
Доступно в on-line версии с 31.12.2014
Познавательные действия человека представляют собой целенаправленные акты, включающие стадию ориентировки в ситуации и предметах действия, а также неотделимую от нее стадию организации и выполнения ответа о свойствах и отношениях изучаемых предметов. На стадии ориентировки осуществляются процессы восприятия или воображения предметов, воспоминание или мышление о них, тогда как на стадии ответа протекают процессы организации и реализации двигательного или речевого ответа о содержании познанного предмета. На ориентировочной стадии познавательного действия могут выполняться не только внутренние психические процессы, но и внешние движения, направленные на ощупывание предмета, на его перемещение или деформацию с целью выяснения его веса, упругости и пр. Артикуляция слов и звуков, воображение различных двигательных актов с каким-либо предметом также могут осуществляться на стадии ориентировки в его свойствах. Нейросетевая модель познавательного действия представляет собой многоуровневую систему (сеть) «математических», формальных нейронов, с помощью которых моделируются механизмы и изучаются принципы работы реальных нейронных сетей головного мозга, обеспечивающих выполнение таких действий. В данной работе излагается нейросетевая модель простых по содержанию и кратковременных познавательных действий, направленных на обнаружение или опознание зрительно воспринимаемых предметов.
Формальный нейрон моделирует некоторые свойства биологического нейрона и является основным функционально-структурным компонентом математической модели нейронной сети (рис. 1). Он состоит из тела (Sk), у которого с одной стороны имеются дендриты, по которым на нейрон через синапсы поступают входные сигналы-воздействия (хi) от других нейронов сети. С другой стороны от тела нейрона отходит аксон, по которому выходной сигнал от нейрона, или его выход (уk), передается на другие нейроны. Каждый входной синапс нейрона характеризуется своим «весом», или синаптическим коэффициентом (wki), который определяет то, с каким знаком и величиной внешнее воздействие (хi) передается на данный нейрон. При wki˃0 синапс является возбуждающим, т.е. положительное воздействие хiна нейрон увеличивает его выход уk, а при wki˂0 —тормозным. Синаптические коэффициенты могут изменяться в зависимости от прошлых входных воздействий (хi) на нейрон, а также в результате обратной связи от его выхода (уk), что позволяет нейронной сети «обучаться на примерах». Приобретаемый человеком опыт познавательных действий также фиксируется в синаптических коэффициентах соответствующих нейронов (перцептивных, моторных и пр.) и в связях между ними.
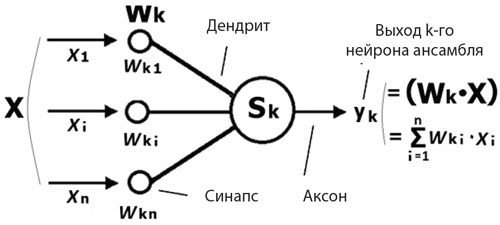
Рис. 1. Формальный нейрон. Sk— тело k-го нейрона ансамбля, X— вектор входного сигнала (х1…xi…хn— его координаты), Wk— вектор синаптических коэффициентов (wk1…wki…wkn— его координаты), уk— выход k-го нейрона, равный скалярному произведению векторов WkиX
Поступающие на синапсы
нейрона внешние воздействия хi, а также «веса»
этих синапсов wki, можно рассматривать
как координаты векторов xи
wk,
расположенных в n-мерном
вещественном линейном пространстве. Базисом этого пространства являются
ортогональные единичные векторы следующего вида: e1=(1,
0…0), e2=(0,
1…0) и т.д. В этом базисе вектор х равен (х1e1+х2e2+…+хnen), а вектор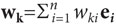 . При этом
величина выхода формального нейрона (уk)
описывается «взвешенной» по синаптическим коэффициентам суммой входных
воздействий[1]:
. При этом
величина выхода формального нейрона (уk)
описывается «взвешенной» по синаптическим коэффициентам суммой входных
воздействий[1]:
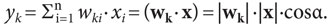
(1)
Формула (1) трактуется также как скалярное произведение векторов (wk٠х), которое равно произведению модулей («длин») этих векторов, умноженному на косинус угла α между ними. Когда cosα=+1, то выход нейрона уk является положительным и максимальным, что бывает при коллинеарности (параллельности) и одинаковой направленности векторов х и wk, т.е. при угле α между ними, равном 0, 360 и т.д. градусов.
Таким образом, на входе формального нейрона имеются два многомерных вектора: вектор x=Σхiei— описывающий состояние внешнего воздействия на нейрон, и вектор wk=Σwkiei— описывающий состояние синаптического входа нейрона, которое «преломляет» внешнее воздействие. В классической модели нейрона его выход описывается вещественным числом уk, равным скалярному произведению векторов xи wk.
В некоторыхмоделях нейрона его выход рассчитывается иначе. В них величина у складывается с порогом срабатывания нейрона (w0), и эта сумма служит аргументом для «передаточной функции» F(у+w0)=р, значение которой определяет выход нейрона. В зависимости от целей построения нейросетевой модели передаточная функция F может быть линейной, сигмоидальной и пр. В рассматриваемой далее сферической нейросетевой модели познавательного действия порог срабатывания нейронов принимается равным нулю, а в качестве передаточной функции берется простейшая тождественная функция, т.е. полагается, что р=у.
Прежде чем перейти к описанию сферической модели нейронной сети, рассмотрим несколько общих принципов структурной организации и функционирования реальных нейронных сетей человеческого мозга, которые учитываются в этой модели и выражаются на математическом языке. К таким принципам относятся следующие: многослойное и иерархическое строение нейронных сетей, топографическое отображение нижних слоев на верхние, «колончатая» организация слоев коры, объединение нейронов в целостные ансамбли, выполняющие различные физиологические и психологические функции.
В разных «отделах» зрительной системы человека, таких как сетчатка глаз, в двух латеральных коленчатых телах (ЛКТ), в первичной зрительной коре и других, нейроны образуют четко различимые слои, в каждом из которых расположены нервные клетки различного вида (Хьюбел, 1990). В сетчатке имеется три слоя нейронов. Первый слой образуют фоторецепторы (палочки и колбочки), которые одним своим отростком (дендритом) обращены к внешнему миру, а другим отростком (аксоном) связаны с дендритами нейронов второго слоя сетчатки, включающим три вида нервных клеток – горизонтальные, амакриновые и биполярные. Третий слой сетчатки образуют ганглиозные клетки, на дендритах которых через синапсы конвергируют аксоны нейронов второго слоя. Аксоны ганглиозных клеток образуют выходящие из сетчатки каждого глаза зрительные нервы, идущие вглубь мозга к нейронам двух ЛКТ. Каждое ЛКТ содержит по 6 нейронных слоев. Большинство нейронов ЛКТ направляют свой аксон в первичную зрительную кору человека, в которой также имеется 6 слоев нейронов (при толщине коры около 2 мм). Благодаря многослойному строению первичную (проекционную) зрительную кору, расположенную в затылочной части мозга и имеющую размеры с почтовую марку, называют также стриарной («полосатой»).
Иерархичность строения нейронных слоев выражается, в частности, в том, что с одной ганглиозной клеткой сетчатки связано несколько нейронов второго слоя, а с каждым из этих нейронов связано несколько рецепторов. Тем самым одна ганглиозная клетка через ряд нейронов второго слоя получает сигналы от нескольких рецепторов, которые образуют рецептивное поле этой клетки. Рецептивное поле одной ганглиозной клетки обладает круговой симметрией и по числу соответствующих ей рецепторов больше, чем рецептивные поля нейронов второго уровня. Иерархичность строения нейронных сетей выражается также в том, что нейроны более высокого уровня отражают своими входными и выходными состояниями более сложные свойства и структуры воздействующего на сетчатку предмета, чем нейроны более низкого уровня. Топографическая организация некоторых нейронных слоев мозга выражается в том, что предшествующая структура проецируется на последующую упорядоченным образом: «если идти вдоль какой-либо линии на сетчатке, то проекции последовательных точек этой линии в ЛКТ и в коре также образуют одну непрерывную линию» (Хьюбел, 1990, с. 65). Кроме того, «если поврежден определенный участок первичной зрительной коры, то развивается “локальная слепота”, как если бы была разрушена соответствующая область сетчатки» (там же, с. 68).
Нейроны из разных слоев первичной зрительной коры, лежащие друг под другом на пересекающем кору перпендикуляре, образуют макро и мини-колонки, по В. Маунткаслу (Эделмен, Маунткасл, 1981). Когда на рецептивное поле одной мини-колонки у кошки или обезьяны Д. Хьюбел и Т. Визел подавали изображение короткой линии, имеющей определенную ориентацию φ (угол между линией и фиксированной прямой), то при движении микроэлектрода перпендикулярно коре и вдоль соответствующей мини-колонки регистрировались потенциалы действия входящих в нее нейронов. При изменении ориентации линии реакция нейронов данной мини-колонки уменьшалась или исчезала совсем. Однако возбуждались нейроны соседней мини-колонки, которые «настроены» на другое значение ориентации линии, предъявляемой в том же рецептивном поле. Поскольку между близлежащими мини-колонками зрительной коры есть горизонтальные тормозные связи, то переход максимального возбуждения от одной колонки на другую при непрерывном изменении ориентации линии φ происходит плавно и может быть описан синусоидой.
При перемещении электрода параллельно поверхности стриарной коры он пересекает ряд мини-колонок, которые реагируют на различные ориентации линии, предъявляемые в некотором участке поля зрения. В стриарной коре приматов одна мини-колонка содержит около 260 связанных по вертикали нейронов, образующих цилиндр диаметром около 30 мкм. Упорядоченные группы мини-колонок образуют макро-колонку, нейроны которой настроены на выделение разных значений некоторого свойства предмета, находящегося в определенном месте поля зрения. Диаметр одной группы мини-колонок, по разному реагирующих на ориентации линий в диапазоне от 0 до 180º, равен примерно 0,5 мм, что соответствует размеру одной макро-колонки. Различные группы специализированных по своим функциям нейронов, расположенные в сенсорной, моторной и ассоциативной коре мозга и имеющие колончатую структуру, в дальнейшем будем называть нейронными ансамблями.
Данное выше краткое и упрощенное описание строения реальной нейронной сети «переводится» далее на математический язык. При этом строится математическая модель того, как нейронная сеть может реагировать на разные значения одномерного свойства предмета, такогокак ориентация линии, направление ее движения и пр., как с помощью этой сети человек управляет движениями, ассоциированными с разными значениями свойства[2]. В такой сети выделяются две взаимосвязанные части — нейронный механизм, определяющий значение свойства объекта (зрительный анализатор), и нейронный механизм организации и осуществления вербального или двигательного ответа, ассоциированного с разными значениями свойства. Взаимодействие между этими частями сети осуществляют нейроны памяти. В опознавательном действии они порождают образы (эталоны) ожидаемых объектов, с которыми соотнесены разные ответы.
В модели зрительного анализатора имеется три слоя нейронов — формальные рецепторы, предетекторы и детекторы (рис. 2). Один слой нейронов в модели может заменять несколько слоев нейронов реальной сети. Так, один слой формальных рецепторов заменяет три слоя реальных нейронов сетчатки. Выходы формальных рецепторов поступают на синапсы предетекторов, под которыми понимаются нейроны, лежащие между рецепторами и детекторами. Предетекторам в излагаемой модели соответствуют нейроны ЛКТ. Выходы предетекторов поступают на синапсы нейронов, расположенных в зрительной коре и образующих ансамбль детекторов. Детекторы настроены и по-разному реагируют на различные значения некоторого свойства предмета, воздействующего на рецепторы.
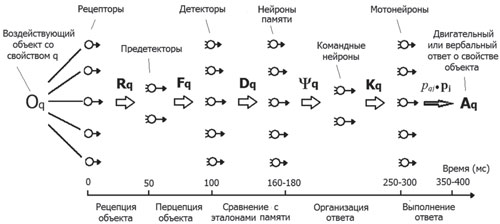
Рис. 2. Схема нейронной сети опознавательного действия. Векторы Rq, Fq, Dq, Yq, Kqописывают состояния объекта со свойством q на выходе соответствующих нейронных ансамблей. Вектор рqj٠рj описывает выход мотонейрона, имеющего максимальное возбуждение в ансамбле и вызывающего внешний ответ на данный объект
Количество предетекторов в сферической модели выбирается на основе данных психологического эксперимента. Оно полагается равным размерности «семантического пространства» того свойства предмета, значения которого должна различать и сравнивать формальная нейронная сеть. Эмпирически размерность этого пространства может определяться с помощью многомерного шкалирования матрицы субъективных оценок попарной близости ряда предметов, различаемых по данному свойству. Так, обработка методом многомерного шкалирования матрицы субъективных оценок близости разных по физической ориентации линий показала, что воспринимаемые линии располагаются на окружности (наодномерной сфере), которая лежит на плоскости, т.е. в двухмерном евклидовом пространстве (Измайлов и др., 2003).
В связи с этим в сферической модели нейронного анализатора различий между линиями разной ориентации имеется два предетектора. Это согласуется с данными Д. Хьюбела, согласно которому для детекции наклона линии достаточно двух предетекторов, реагирующих на противоположные концы линии. Возбуждения двух предетекторов трактуются в модели как координаты фиксированного по длине радиус-вектора F (см. далее), который своим концом указывает положение предмета на окружности в семантическом пространстве, построенном по субъективным оценкам близостей разных предметов, различаемых по одномерному свойству.
Аналогично этому размерность семантического пространства цветовых стимулов у человека равна четырем, поскольку при их многомерном шкалировании они располагаются в четырехмерном пространстве и лежат на поверхности трехмерной сферы, образующей трехмерное подпространство в этом пространстве (Измайлов и др., 1989). Видимый цвет как психологическое свойство предметов воспринимается человеком с помощью 4 нейронов-предетекторов (красно-зеленого, сине-желтого, яркостного и темнового), соответствующих четырем осям семантического пространства цветов, между которыми имеется одна геометрическая связь, определяющая длину радиус-вектора трехмерной сферы, на поверхности которой расположены шкалируемые цветовые стимулы.
Поэтому в сферической модели цветового зрения человека цвет стимула трактуется как его трехмерное свойство, которое описывается тремя числами — сферическими координатами радиус-вектора постоянной длины, направленного в определенную точку сферы, в которой находится данный цвет и соответствующий ему формальный нейрон-детектор. При этом субъективно близким цветам соответствуют пространственно близкие точки на математической сфере, расположенной в четырехмерном пространстве, а также близкие точки на проекциях этой сферы на разные плоскости. В связи с этим нейросетевые модели, объясняющие расположение на поверхности n-мерной сферы (n≥1) различных предметов, субъективная близость которых шкалируется по какому-либо n-мерному свойству, называются сферическими.
В сферической модели
нейронной сети объективное состояние воздействующего на органы чувств объекта
со свойством jq(где
j — название свойства
объекта, а q — номер его значения) описывается вектором Xq, который поступает на
синаптический вход Wkкаждого
k-го рецептора (нейрона) и образует с ним скалярное
произведение, равное числу 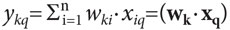 (см.
рис. 1 и 2). Числа ykq описывают выход k-го
рецептора при воздействии на него воздействия Xq. Каждому такому числу
может соответствовать некоторая величина возбуждения нейрона сетчатки или
волоскового рецептора уха, настроенного на определенную частоту акустического
спектра.
(см.
рис. 1 и 2). Числа ykq описывают выход k-го
рецептора при воздействии на него воздействия Xq. Каждому такому числу
может соответствовать некоторая величина возбуждения нейрона сетчатки или
волоскового рецептора уха, настроенного на определенную частоту акустического
спектра.
На выходе ансамбля рецепторов,
участвующих в восприятии объекта, состояние этого объекта уже является
физиологическим и описывается вектором Rq с координатами {уiq}.
Такая трансформация объективного состояния воздействующего на сетчатку объекта
со свойством jq в его физиологическое
состояние в излагаемой модели описывается «оператором» (точнее, линейным
отображением) Wф,
преобразующим вектор Xqв
вектор Rq,
в котором сохранена информация о свойстве объекта jq. Этот оператор описывает
переход объекта из одного состояния в другое: Rq=Wф(Xq). Он может служить
моделью процессафизиологического
отражения объекта, протекающего на уровне рецепторов нейронной сети. В
координатном представлении оператору Wф соответствует
матрица, содержащая m
строк (по числу рецепторов в ансамбле) и n столбцов, где n — число синапсов у одного
рецептора (см. рис. 1). Векторы Xq и Rq представлены столбцами
своих координат. Действию оператора Wф на
вектор Xq
соответствует умножение матрицы этого оператора на вектор-столбец Xq. В строках матрицы Wф
стоят синаптические коэффициенты каждого из m рецепторов ансамбля. При
этом выход k-го
рецептора, или k-я
координата вектора Rq,
вычисляется по правилам матричного умножения и равна: 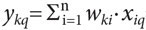 , что совпадает с формулой 1.
, что совпадает с формулой 1.
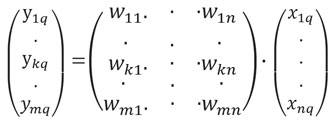
Возбуждение ансамбля рецепторов, т.е. его выходной вектор Rq, поступает далее на синаптические входы Cjкаждого нейрона-предетектора, где преобразуется по формуле (2). В результате выход j-го предетектора возбуждается до величины[3]:
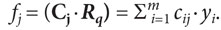
(2)
Формула (2) аналогична формуле (1), поскольку каждое число fjпредставляет собой скалярное произведение вектора синаптических коэффициентов j-го предетектора Cj=(сj1, сj2, … сjm) на входящий в этот ансамбль вектор Rq=(у1, у2, … уm). Числа сjiравны синаптическим коэффициентам i-го синапса у j-го предетектора, а числа fjобразуют координаты вектора Fq, описывающего возбуждение, или выходное состояние всего ансамбля предетекторов при воздействии на сетчатку объекта в состоянии Xq.
В сферических нейросетевых моделях познавательных процессов (Соколов, Вайткявичус, 1989; Фомин и др., 1979) предполагается, что в достаточно развитых нейронных сетях возникают такие ансамбли предетекторов, у которых выходные векторы Fявляются «нормированными», т.е. единичными по длине в евклидовой метрике[4]. Из этого следует, что значения свойства jqна выходе таких ансамблей нейронов кодируются не длиной вектора их возбуждения Fq(которая в данном случае равна единице и не зависит от jq), а его направлением, которое определяется углами ajэтого вектора с осями координат. При этом координаты единичного вектора Fq, равные его проекциям на «координатные оси», будут равны: fj=cos(aj(jq)), где aj—зависящий от jq угол между вектором Fqи j-й координатной осью.
Наиболее простой зависимостью углов aj от φq является линейная, согласно которой aj(jq)=ljq+jjo, где l — константа, одинаковая для всех aj. В этом случае координаты единичного вектора Fqравны: fj=cos(ljq+jjo), где jjo— фазы координат. Как показано выше, для кодирования значений jq одномерного свойства требуется ансамбль из двух предетекторов, порождающий двухмерный вектор Fq. Отношение между фазами координат этого вектора должно быть таким, чтобы выполнялось условие единичности его длины, т.е. модуль |Fq|=f12+f22=1. С помощью простых тригонометрических выкладок можно показать, что это условие выполняется тогда, когда фазы координат вектора Fqсвязаны равенством j1o=90º‒j2o. Если фазу одной координаты, например первой, выбрать равной нулю, т.е. взять f1(jq)=cos(ljq),то фаза другой координаты f2(jq)будет равна 90º, т.е. эта координата будет равна f2(jq)=cos(ljq+90º), что в свою очередь равно sin(ljq). (Произвольный выбор значения одной из фаз возможен, если при этом не нарушается условие единичности вектора Fq.) Таким образом, координаты вектора возбуждения Fqансамбля из двух предетекторов являются гармоническими функциями от значений jq одномерного свойства стимула, а сам единичный по длине вектор Fq, кодирующий состояние предмета со свойством φq на выходе ансамбля предетекторов, равен:
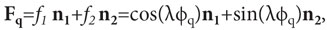
(3)
где n1 и n2— ортогональные базисные векторы пространства, в котором расположены различные векторы F.
Константа l в формуле (3) связана со способностью человека к различению разных значений свойства j, а также с порогами чувствительности к этому свойству. Так, угол между двумя векторами F(j1) и F(j2), кодирующими разные значения свойства j, равен l(j2‒j1). При этом, как показано Е.Н. Соколовым и Ч.А. Измайловым, чем больше угол между двумя векторами Fи пропорциональная этому углу длина хорды или дуги между концами этих векторов, тем больше субъективные различия между кодируемыми ими значениями свойства. Поскольку при возрастании l и фиксированном Δjугол между векторами F(j1) и F(j2) возрастает, то субъективное различие между значениями свойства j1 и j2 также увеличивается, что означает повышение чувствительности к этому свойству.
Таким образом, формула (3) каждому значению свойства φqсопоставляет пару взаимосвязанных координат вектора Fq. Эти координаты можно трактовать как «кодовое слово», которым в модели нейронной сети кодируется значение одномерного свойства. Само же преобразование свойства φq в данное кодовое слово и в соответствующий ему вектор Fq можно рассматривать как векторное кодирование в нейронной сети данного значения свойства. Иначе говоря, векторное кодирование свойства jq в нейронной сети состоит в том, что оно с помощью различных преобразований, описываемых математическими функциями, преобразуется в координаты выходных векторов разных нейронных ансамблей.
Возникает вопрос о том, как в нейронной сети осуществляется обратное преобразование, т.е. операция декодирования векторов сети путем отображения их в соответствующие значения свойств стимула. Как, например, на основе декодирования имеющегося в сети вектора Fqможет определяться соответствующее ему значение свойства φq?В сферической модели этот вопрос решается следующим образом. Полагается, что декодирование значения свойства может осуществляться с помощью расположенных в коре мозга нейронов-детекторов, на синаптические входы которых поступает вектор Fq(см. рис. 2). Каждый k-й детектор характеризуется своим входным состоянием, описываемым вектором синаптических коэффициентов Pk с координатами {pkj}. При этом значение выхода dkкаждого детектора равно скалярному произведению векторов Pk и Fqи определяется по формуле (4), аналогичной формулам 1 и 2:
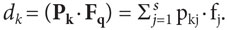
(4)
В сферической нейросетевой модели предполагается, что при воздействии на рецепторы предмета со свойством j нейронная сеть определяет его значение jqпо номеру q или по нумерованному месту на «детекторном экране» того нейрона, который имеет максимальное возбуждение при данном воздействии (Фомин и др., 1979). Таким детектором является тот, у которого вектор синаптических коэффициентов Pkсовпадает с вектором возбуждения Fqвсего ансамбля предетекторов, т.е. имеет вид: Pq=cos(ljq)n1+sin(ljq)n2. В этом случае единичные векторы Fqи Pqявляются параллельными и однонаправленными, а их скалярное произведение, описывающее выход q-го детектора, является максимальным и равно единице: (Fq٠Pq)=dq=1. Максимальное значение выхода q-го детектора является сигналом, по которому мозг может определить, что на рецепторы воздействует значение свойства jq, имеющее такой же номер, как и номер соответствующего детектора.
Входные векторы детекторов, соседних с максимально возбужденным, имеют другие координаты, например {cos(ljg), sin(ljg)}, где g≠q. Они настроены на выделение других значений jg свойства j и не параллельны вектору Fq. Поэтому скалярные произведения векторов Pg и Fq меньше единицы, а выходные возбуждения детекторов с номерами gне являются максимальными в данном ансамбле. Такое представление о механизме определения свойства jq согласуется с результатами экспериментов Хьюбела и Визела, предъявлявших на одно и то же место сетчатки линии разной ориентации jq и регистрировавших возбуждения близко лежащих нейронов коры. Разным значениям свойства jqсоответствовали разные по пространственной локализации максимально возбужденные нейроны данного участка коры.
Таким образом, выход максимально возбужденного q-го детектора первичной зрительной коры описывается вектором dq٠mq, где mq—
единичный базисный вектор линейного пространства, в котором расположен вектор D, описывающий выход всего ансамбля
детекторов. Выделение мозгом максимального по длине вектора dq٠mqможно
рассматривать как «редукцию» вектора D,
который равен сумме выходных векторов всех детекторов ансамбля, число которых
равно v: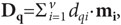 ,
где dqi—
возбуждение i-го детектора при воздействии на его
вход вектора Fq.
,
где dqi—
возбуждение i-го детектора при воздействии на его
вход вектора Fq.
Вектор Dq, как и вектор Fq, также несет в себе информацию о значении свойства jq, кодируя его своими углами с векторами mi, образующими ортогональный базис пространства векторов D. При редукции вектора Dqпутем его сведения к вектору dq٠mqчасть информации о свойстве jq теряется, что не позволяет последнему вектору быть полноценной основой для быстрой организации внешнего двигательного или вербального ответа на это свойство. Для выполнения таких ответов на ансамбль командных нейронов, имеющих большое число (v) синапсов, должен поступать «полный» вектор Dq с такой же размерностьюv, а не выделенная из него (редуцированная)компонента dq٠mq. Однако если на стимулыне нужно давать внешний двигательный ответ, то процессы выделения мозгом максимально возбужденных нейронов на уровне детекторов зрительной коры и нейронов памяти, т. е. процессы редукции векторов Dq и Yq (см. рис. 2) могут, по-видимому, играть важную роль в процессе осознания человеком ощущений разных значений свойства j.
В соответствии с разработанной А.Н. Леонтьевым и А.В. Запорожцем деятельностной концепцией возникновения ощущений (Леонтьев, 1959) можно предположить, что воздействующее на рецепторы значение свойства jq становится неосознанно ощущаемым, когда результат его отражения на некотором уровне нейронной сети начинает участвовать в регуляции и направлять ассоциированное с ним внешнее движение субъекта. Этому условию удовлетворяет вектор Dq, который начинает участвовать в регуляции движения тогда, когда поступает на входы нейронов памяти (эталонов), с которыми ассоциированы различные «ответные» движения на разные значения свойства j.
Векторы Fqи Dqможно рассматривать как математические модели физиологического и психологического (ощущаемого) состояний предмета, воздействующего на человека и его мозг. Такая интерпретация этих векторов согласуется с общенаучным векторным описанием различных состояний и основана на деятельностном представлении о процессе отражения предмета, как совокупности преобразований его исходного объективного состояния в различные физиологические и/или психологические состояния (Беспалов, 2008).
Входящие в формулу 4 числа рji образуют матрицу (рji) преобразования Р, которое переводит вектор Fqв вектор Dq=Р٠Fq. Преобразование Р можно рассматривать как математическую модель той части познавательного процесса, которая соответствует переходу воспринимаемого предмета из состояния Fqв Dq. Вместе с тем саму матрицу (рji), строки которой составлены из координат векторов синаптических коэффициентов детекторов, можно трактовать как модель перцептивного средства, с помощью которого воспринимаемый предмет из физиологического состояния Fqпреобразуется в неосознанноощущаемое состояние Dq. Это состояние предмета может затем осознаваться с помощью ансамблей нейронов на более высоких уровнях сети.
Возможный психофизиологический механизм осознания ощущения описан А.М. Иваницким (1996) в концепции «синтеза информации о физических и сигнальных свойствах стимула на нейронах проекционной коры», а также Дж. Эделменом (Эделмен, Маунткасл, 1987) в концепции «повторного входа» возбуждения в зрительную кору. Согласно концепции А.М. Иваницкого, первичное возбуждение проекционной (первичной) зрительной коры возникает через 50 мс после показа стимула. Далее оно перемещается в височную и лобную кору, а затем от височной области в гиппокамп и гипоталамус. После этого возбуждение вновь возвращается к проекционной коре и повторно ее активирует. При прохождении по разным отделам мозга первичное возбуждение обогащается прошлым опытом и хранящейся в памяти информацией о стимуле, о текущих потребностях человека и пр. В результате первичное возбуждение зрительной коры (в нашей модели вектор Dq и соответствующее ему неосознанное ощущение) через 160—180 мс после показа стимула превращается в осознанное ощущение стимула. Данную концепцию А.М. Иваницкий обосновывает, в частности, результатами экспериментов с регистрацией вызванных потенциалов (ВП).
В наших экспериментах (Беспалов, Вартанов, 2014) при показе испытуемому 4 одиночных стимулов Струпа (слов «зеленый» или «красный», написанных зеленым или красным шрифтом) на записи ВП также наблюдается небольшой позитивный пик активности (Р50) на латенции 50 мс в затылочных О1 и О2 отведениях ЭЭГ и в близких к ним латеральных отведениях Т5 и Т6. Этот пик имеет амплитуду около +3 мкВ, которая на 1,5 мкВ выше фоновой. Затем происходит резкое (за 50 мс) уменьшение амплитуды этих ВП до отрицательных значений (от –6 до –8 мкВ), после чего амплитуда ВП снова увеличивается на 4—10 мкВ (в зависимости от отведения). В результате в указанных отведениях возникают резкие негативные пики ВП (N100) на латенции 100 мс. Такая картина поведения ВП наблюдается при решении задач, в которых испытуемый читал слово «про себя» сразу после его предъявления на экране, называл «про себя» цвет шрифта слова, а также в задаче, требующей осознавать появление стимулов на экране при их пассивном восприятии, без целенаправленного чтения или называния цвета шрифта.
Первый позитивный пик Р50 в парных отведениях Т (4,5) и О можно, по-видимому, соотнести с возбуждением ансамбля предетекторов (в ЛКТ), которое через 50 мс после показа стимула поступает на 4-й (входной) слой ансамбля детекторов первичной зрительной коры. На латенции 100 мс аналогичный позитивный пик ВП (Р100) наблюдается уже в височных (Т3 и Т4) и лобных (F) отведениях ЭЭГ, что может свидетельствовать о перемещении возбуждения в эти области мозга. После этого в этих отведениях также происходит резкое (за 50—80 мс) уменьшение амплитуды этих ВП до отрицательных значений (около 8 мкВ). В результате в височных и лобных отведениях тоже возникают резкие негативные пики ВП на латенциях от 150 до 180 мс, которые наблюдаются также в теменных отведениях (P) и в области центральной борозды (С).
Возможно, что первый негативный пик ВП в затылочной области коры (N100 в отведениях Т (4,5) и О) отражает возникающее в ней возбуждение, описываемое вектором Dq, а также соответствующее ему неосознанное зрительное ощущение значения свойства jq(цвета и конфигурации букв в слове). Это ощущение затем осмысливается, путем его соотнесения с опознавательными эталонами и осознается (путем выражения в языке и соотнесения с образом Я) с помощью нейронов в других отделах мозга. Процессам осмысления ощущения могут соответствовать резкие негативные пики ВП в других отведениях ЭЭГ (F, C, P, T3 и T4) на латенциях от 150 до 180 мс, которые можно, по-видимому, трактовать как выражение процесса соотнесения ощущения свойства объекта с опознавательными эталонами.
В результате соотнесения неосознанного ощущения с хранящимися в памяти опознавательными эталонами вектор Dq преобразуется в вектор Yq, который описывает состояние распределенного по мозгу ансамбля взаимосвязанных нейронов-детекторов, участвующих в осмысленном восприятии свойства предмета. Такие нейроны Е.Н. Соколов (2004) относил к категории «нейронов сознания». Согласно ему, «в основе субъективных явлений лежит активность “нейронов сознания”, образующих распределенную субсистему. С этой точки зрения поток сознания может быть представлен сменяющими друг друга возбуждениями “нейронов сознания”» (Соколов, 2010, с. 115). «…Они составляют отдельный класс нейронов коры, каждый из которых связан с отдельным психическим актом. Поэтому выпадение отдельного “нейрона сознания” ведет к утрате связанного с ним психического переживания» (там же, с. 132). «Особенностью “нейронов сознания” является их зависимость от нейронов активирующей и инактивирующей систем» (там же, с. 118). В связи с этим различные формы «отключения сознания» (при обмороке, в некоторых фазах сна и пр.), а также механизмы «восстановления сознания» после черепно-мозговой травмы Е.Н. Соколов соотносил с изменением режима работы «нейронов сознания», необходимый уровень активации которых поддерживается ретикулярной системой.
Таким образом, вектор Yqописывает выходное состояние распределенного по мозгу ансамбля нейронов, который участвует в отражении q-го значения свойства предмета в форме его осмысленного (частично осознанного) ощущения. Ансамбль нейронов, порождающий вектор Yq, можно рассматривать как динамичную функциональную систему, которая выполняет функцию осмысления первичных, не осознанных ощущений значений свойства φ путем их соотнесения с опознавательными эталонами, которым соответствуют разные ответные движения. Этот ансамбль формируется в мозге из имеющихся нейрональных ресурсов тогда, когда у человека возникает осознанная цель воспринимать разные значения свойства предмета для того, чтобы выполнить по отношению к ним какие-либо акты — назвать эти значения свойства, нажимать соответствующие им кнопки и пр. При формировании такой цели некоторое значение свойства предмета приобретает для человека определенное «сигнальное значение» (отношение к определенному движению) и операциональный смысл (отношение к эталонам, как средствам познавательного действия)[5].
Когда же значения свойства предмета (его положение в пространстве, форма и пр.) изменяются быстро и неожиданно для человека, то они могут отражаться неосознанно и без заранее поставленной цели. В этом случае выход ансамбля детекторов первичной зрительной коры, т.е. вектор D, может сразу (минуя нейроны памяти) поступать на командные нейроны и вызывать определенный двигательный акт, например рефлекторное подхватывание рукой случайно выпавшего из нее предмета. При этом осознанные ощущения неожиданно изменившихся значений свойств предмета будут возникать с помощью «нейронов сознания» уже после осуществления рефлекторного двигательного акта.
Отличительной особенностью развиваемой здесь нейросетевой и одновременно деятельностной концепции порождения осмысленных ощущений является утверждение о том, что они возникают с помощью, или посредством, мозга и его нейронных функциональных систем, но существуют и локализуются не в мозге (Леонтьев, 2004), а в перцептивном пространстве субъекта (человека или животного). Такое пространство отличается от физического и представляет собой упорядоченное разными отношениями (ближе/дальше и пр.) множество чувственно воспринимаемых и внешних по отношению к мозгу вещей, на воспринимаемой поверхности которых локализуются ощущения их свойств. Ощущения — это дискретные продукты («кванты») психологического взаимодействия человека с миром, осуществляемого с помощью нейронных ансамблей мозга, работа которых в излагаемой модели описывается операторами, преобразующими объективное состояние воздействующего на нейроны внешнего предмета в его различные психологические состояния, которые отражают этот предмет и представлены в форме его ощущений и образов.
После того как q-е значение свойства предмета психологически отражено в выходных состояниях Dq или Yqсоответствующих нейронных ансамблей мозга, может осуществиться ассоциированное с этим значением рефлекторное или произвольное движение или процесс его названия словом. Для описания этих процессов в сферической нейросетевой модели познавательного действия предполагается, что векторы Dq или Yqпосле их нормировки (приведения к единичной длине) поступают на входы ансамбля командных нейронов, который работает по описанным выше принципам.
Если отражаемое свойство предмета является одномерным, то ансамбль командных нейронов содержит два нейрона, как и ансамбль предетекторов. Когда на входы командных нейронов поступает вектор Y, то на выходе этого ансамбля возникает двухмерный единичный вектор Кq=К٠Yq, где К — преобразование, матрица которого имеет 2 строки, в которых расположены синаптические коэффициенты двух командных нейронов и s столбцов — по числу нейронов в ансамбле Yq. Вектор Кq=к1٠к1+к2٠к2 симметричен вектору Fq (см. рис. 2) и имеет аналогичные ему координаты к1=cos(β٠φq) и к2=sin(β٠φq). Он продолжает в новой, моторной форме кодировать воздействующее на рецепторы значение свойства φq. Константа β, входящая в координаты выходного состояния ансамбля командных нейронов, аналогична константе λ в уравнении 3. Она связана с дифференциальной чувствительностью человека к выполняемым движениям, ассоциированным с разными значениями φq. Чем больше величина β, тем на больший угол (β٠Δφq) изменяется направление вектора Кqпри фиксированном изменении (Δφq) параметра φq. Вместе с тем угол между двумя векторами К коррелирует с величиной субъективных различий между соответствующими им движениями. Поэтому чем больше константа β, тем лучше субъект различает две моторные команды, управляющие движениями, ассоциированными с двумя близкими значениями свойства φ.
Вектор Кq поступает на входы мотонейронов,
управляющих разными группами мышц, участвующими в выполнении определенного
двигательного или вербального ответа Аq. Каждый
моторный нейрон в результате формирования условно рефлекторных или смысловых связей
между стимулами и реакциями может быть
настроен на определенное значение jj свойства стимула.
Такая настройка осуществляется путем формирования у j-го мотонейрона входного единичного вектора Mj,
кодирующего соответствующие значения стимульного свойства jj. Координатами векторов
Mjявляются
синаптические коэффициенты соответствующих мотонейронов. Скалярное произведение векторов Кq и Mjопределяет
выход рqj у
j-го
мотонейрона при воздействии на него вектора Кq: (рqj=Кq٠Mj). Каждое число рqjявляется
координатой выходного вектора (рqj٠рj) j-го мотонейрона, где рj—
единичный выходной вектор этого нейрона. При этом выходное состояние всего ансамбля мотонейронов при воздействии на
рецепторы свойства φqописывается
вектором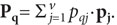 .
.
В сферической нейросетевой модели исполнительной части познавательного действия предполагается, что в ответ на объект со свойством φq выполняется то движение, которое регулируется мотонейроном, получившим максимальное возбуждение при воздействии на него вектора Кq. Таким нейроном является тот, чей вход заранее настроен на φq и описывается вектором Mq, имеющим те же координаты, что и Кq. В этом случае скалярное произведение указанных векторов будет максимальным и равным единице: (Mq٠Кq)=рqq=1. Конкретный словесный или двигательный ответ, ассоциированный со свойством φq и выполняемый при максимальном возбуждении q-го мотонейрона, можно трактовать как внешнюю форму декодирования («редукции») многомерного вектора Рq, а также как внешнюю форму выражения воспринятого человеком значения свойства φq в виде нажатой кнопки, произнесенного слова и пр.
Таким образом, после двигательного (практического) ответа на воспринятое свойство φq оно получает для совершившего его человека свое внешнее «инобытие» в форме определенного следа, или результата преобразования им внешнего материального мира. Полученный результат действия может восприниматься тем же человеком и соотносится им с образом желаемого или планируемого результата, что происходит при оценивании правильности своего ответа или при сравнении заданного и полученного результата действия. Эти процессы обратного соотнесения своего ответа с обусловившим его воздействием φq и с целью действия входят в это же действие и выполняются в нем на стадии восприятия, осознания и коррекции полученного результата. Однако описание этих процессов с позиций изложенной выше сферической нейросетевой модели познавательного действия сталкивается с трудностями, которые обусловлены отсутствием в этой модели обратных связей, а также описания механизма сравнения полученного и заданного результата, оценивания правильности ответа и пр.
В настоящее время в работах Е.Н. Соколова и его соратников (Г.Г. Вайткявичуса, А.В. Вартанова, Ч.А. Измайлова, Г.В. Лосика, А.М. Черноризова и др.) разработаны сферические модели цветового зрения, модели стереоанализатора и анализатора интенсивности стимула, модель восприятия речевых стимулов и др. Раскрыты нейронные механизмы ориентировочного и условного рефлексов, константности восприятия, маскировки стимула и возникновения некоторых зрительных иллюзий. Учет латерального торможения между детекторами в сферической модели позволил описать механизм увеличения дифференциальной чувствительности анализатора при длительном наблюдении адаптирующего стимула, механизм описанного Дж. Гибсоном «эффекта нормализации» наклонной линии. Как показывают исследования, сферические модели познавательных процессов хорошо согласуется с результатами многомерного шкалирования субъективных и объективных (по амплитудам ВП) оценок близости предметов, сравниваемых по одномерному или многомерному свойству.
Вместе с тем в сферических нейросетевых моделях отсутствуют математические средства для описания динамики и длительности познавательных процессов. Эти модели в большей степени описывают результаты психических процессов, например результаты оценивания близости предметов по некоторому свойству, но они не являются моделями самих процессов сравнения этих предметов. Статичные представления о работе нейронных ансамблей не позволяют строить модели обнаружения и опознания предметов в хронометрических экспериментах, которые объясняли бы зависимость времени опознания предметов от условий их восприятия и способов организации ответа. Это связано с тем, что в сферической модели отсутствует временной параметр, не вычисляются вероятности правильного и неправильных ответов на воздействие φq, не описывается возможный механизм взаимовлияния («интерференции») различных процессов решения познавательных задач.
Для устранения этих недостатков в сферическую модель познавательного процесса было введено время и произведен переход в область комплексных чисел, описывающих координаты векторов F и D (Беспалов, 2010). В получаемой таким образом квантово-подобной модели познавательного процесса, использующей тот же аппарат линейной алгебры, что в квантовой физике, появляется возможность математического (но при этом не физического, а психологического) описания возможных механизмов осознания ощущений разных значений свойств предмета, механизма интерференции психологических процессов и пр. Более детально эту модель познавательного процесса и вытекающие из нее философские, теоретические и экспериментально проверяемые следствия планируется изложить в следующей статье на данную тему.
Список литературы
Беспалов Б.И. Классический и неклассический подход к теоретическому описанию психических состояний человека // Психология психических состояний: теория и практика. Мат-лы 1-й Всеросс. науч.-практ. конф. (Казан. гос. ун-т, 13—15 ноября 2008 г.) Ч. I. Казань: ЗАО «Новое знание», 2008. С. 112—115.
Беспалов Б.И. Психофизиологическая проблема в деятельностной психологии человека // Мат-лы 4-й Междунар. конф. по когнитивной науке (Томск, 22—26 июня 2010 г.). Томск, 2010. Т. 1. С. 171—173. URL: http://www.psy.msu.ru/people/bespalov/bespalov_2010.pdf
Беспалов Б.И. Развитие представлений П.Я. Гальперна о средствах, орудиях и орудийных операциях человека и животного // Культурно-историческая психология. 2012. № 4. С. 55—66. URL: http://www.psy.msu.ru/people/bespalov/bespalov_2012a.pdf
Беспалов Б.И., Вартанов А.В. Вызванные потенциалы как показатели интерференции психических процессов в задаче Струпа // Нейронаука для медицины и психологии: Труды 10-го Международного междисциплинарного конгресса (Судак, 2—12 июня 2014 г.). М.: МАКС Пресс, 2014. С. 82—83.
Иваницкий А.М. Мозговая основа субъективных переживаний: гипотеза информационного синтеза // Журнал высш. нерв. деят. им. И.П. Павлова. 1996. Т. 46. № 2. C. 241—252.
Измайлов Ч.А., Соколов Е.Н., Черноризов А.М. Психофизиология цветового зрения. М.: Изд-во Моск. ун-та, 1989.
Измайлов Ч.А., Соколов Е.Н., Коршунова С.Г., Чудина Ю.А. Геометрическая модель различения ориентаций линии, основанная на субъективных оценках и зрительных вызванных потенциалах // Журнал высш. нерв. деят. им. И.П. Павлова. 2003. Т. 54. № 2. С. 267—279.
Леонтьев А.Н. Деятельность. Сознание. Личность. М.: Смысл, 2004.
Леонтьев А.Н. Проблемы развития психики. М.: Изд-во Академии пед. наук РСФСР, 1959.
Соколов Е.Н. Восприятие и условный рефлекс: Новый взгляд. М.: УМК «Психология», 2003.
Соколов Е.Н. Нейроны сознания // Психология. Журнал высшей школы экономики. 2004. Т. 1. № 2. С. 3—15.
Соколов Е.Н. Очерки по психофизиологии сознания. М.: Изд-во Моск. ун-та, 2010.
Соколов Е.Н., Вайткявичус Г.Г. Нейроинтеллект: От нейрона к нейрокомпьютеру. М.: Наука, 1989.
Фомин С.В., Соколов Е.Н., Вайткявичус Г.Г. Искусственные органы чувств. Моделирование сенсорных систем. М.: Наука, 1979.
Хьюбел Д. Глаз, мозг, зрение. М.: Мир, 1990.
Эделмен Дж., Маунткасл В. Разумный мозг. М.: Мир, 1981.
Примечания.
1. Векторы х,wи др. выделяются в текстежирным шрифтом, а их координаты (хi, wiи др.), представляющие собой проекции этих векторов на базисные векторы ei, пишутся курсивом.
2. Под одномерным свойством здесь понимается такое свойство предмета, значения которого могут быть выражены одним вещественным числом. Так, разные значения ориентации (одномерного свойства) линии на плоскости описываются величиной угла между произвольной и эталонной линиями, а для описания цвета линии (ее трехмерного свойства) требуются три числа, например значения цветовых координат RGB.
3. Для упрощения формул индекс q, кодирующий значение свойства отражаемого объекта, в обозначениях координат fj, yiи др. далее не ставится. Однако он сохраняется в обозначениях векторовFq, Rqи др., описывающих отраженное состояние и «образ» объекта со свойством jqна разных уровнях нейронной сети мозга.
4. В евклидовой метрике длина («норма») вектора равна квадратному корню из суммы квадратов его координат. На более низких уровнях нейронной сети мозга длина выходного вектора некоторых нейронных ансамблей (например, в сетчатке) может вычисляться в «сити-блок» метрике, в которой она равна сумме модулей его координат. Возможные физиологические механизмы таких «нормировок» векторов в нервной системе человека рассматриваются в работах Е.Н. Соколова (2010, с. 85, 175 и др.).
5. О понятиях «операциональный смысл» и «средство активности» см. в работе Б.И. Беспалова (2012).
Для цитирования статьи:
Беспалов Б.И. Сферическая нейросетевая модель познавательного действия// Вестник Московского университета.Серия 14.Психология.- 2014.- №4 -с.56-75.












